|
|
|
|
 |
 |
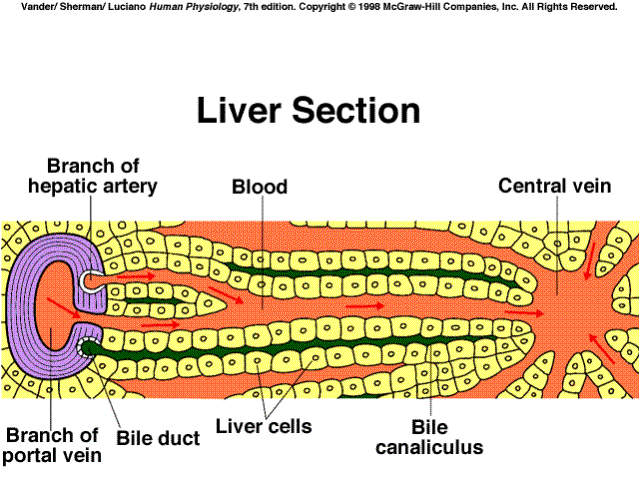 |
The liver has a wide variety of functions, among these are:
|
|
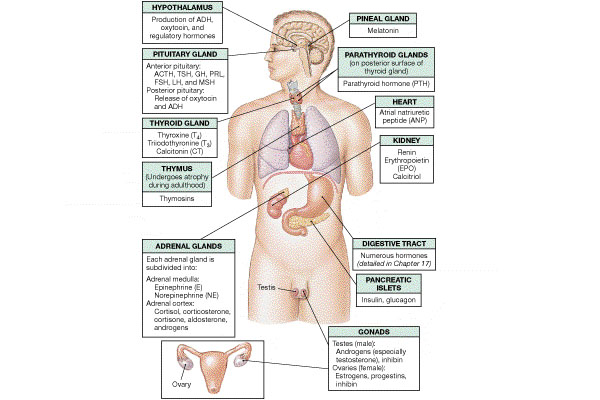 |
Synthesizes and secretes plasma albumin, acute phase proteins, binding proteins for steroid hormones, lipoproteins, and other proteins mentioned later on this page
|
|
 |
|
|
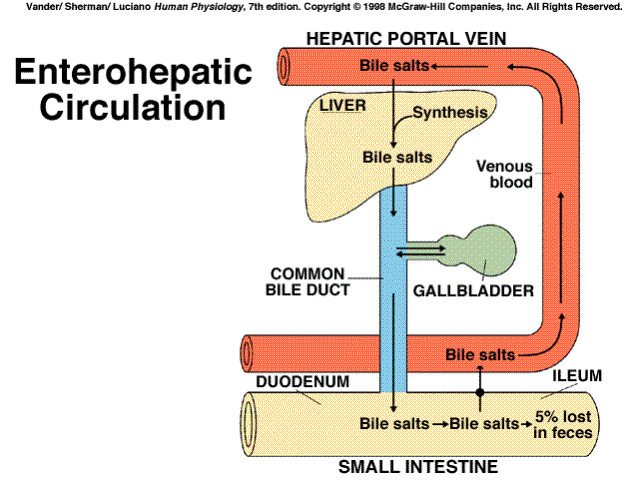 |

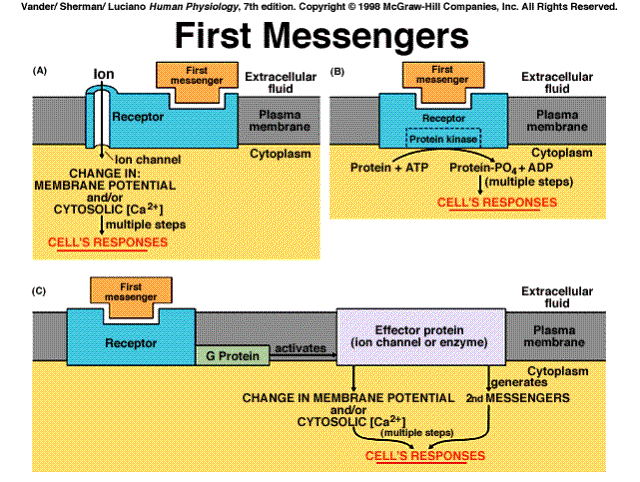
Messenger: That which is used to initiate a chain of events at a receptor site distant from the origin.
|
|
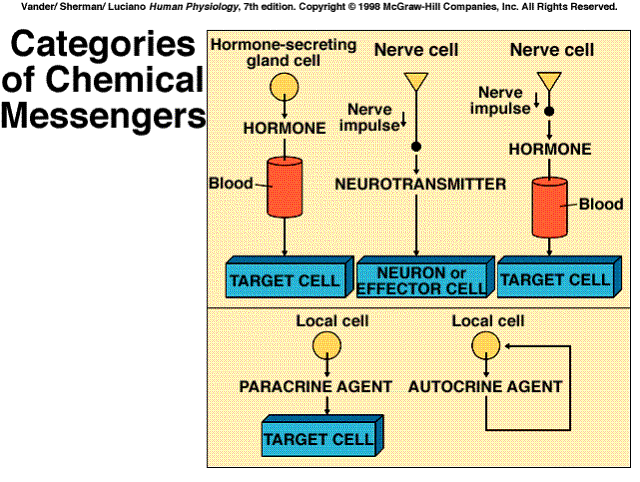 |
Receptor: A specific protein in either the plasma membrane or interior of a target cell with which a chemical messenger combines
Specificity: The ability of a receptor to bind only one type or a limited number of structurally related types of chemical messengers
|
|
 |
Saturation: The degree to which receptors are occupied by a messenger. If all are occupied, the receptors are fully saturated; if half are occupied, the saturation is 50 percent, and so on
Affinity: The strength with which a chemical messenger binds to its receptor
Competition: The ability of different molecules very similar in structure to combine with the same receptor
Antagonist: A molecule that competes for a receptor with a chemical messenger normally present in the body. The antagonist binds to the receptor but does not trigger the cell's response
Agonist: A chemical messenger that binds toa receptor and triggers the cell's response; often refers to a drug that mimics a normal messenger's action
Down-regulation: A decrease in the total number of target-cell receptors for a given messenger in response to chronic high extracellular concentration of the messenger
Up-regulation: An increase in the total number of target-cell receptors for a given messenger in response to a chronic low exracellular concentration of the messenger
Supersensitivity: The increased responsiveness of a target cell to a ginven messenger, resulting from up-regulation
|
|
 |
|
|
 |
|
|
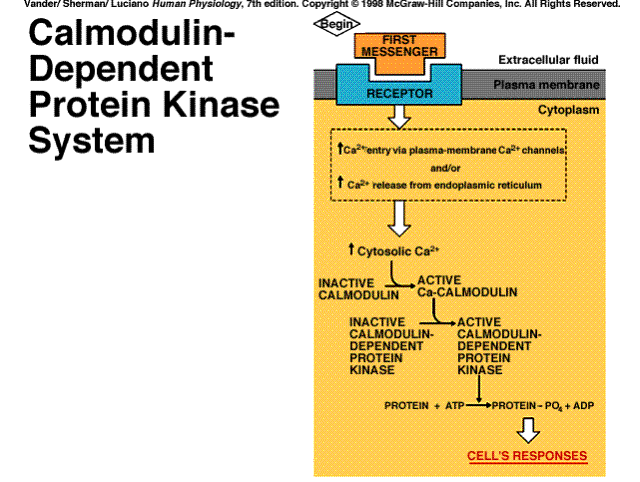 |
+source: Enters cell through plasma membrane ion channels or is released from endoplasmic reticulum
+effects: Activates calmodulin and other calcium binding proteins; calcium-calmodulin activates calmodulin-dependent protein kinases
|
|
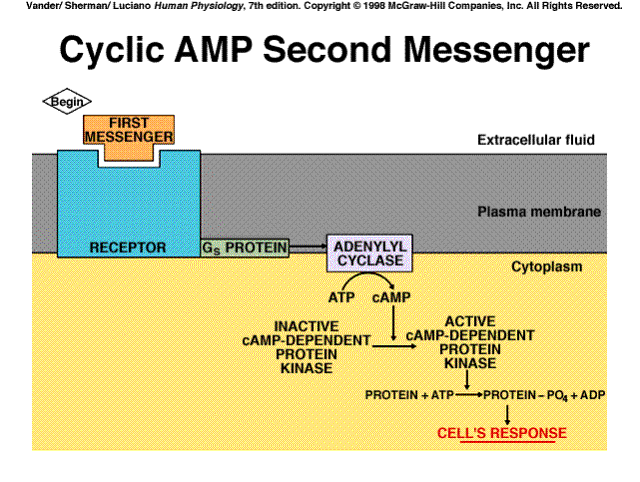 |
+source: A G protein activates plasma membrane adenylyl cyclase, which catalyzes formation of cAMP from ATP
|
|
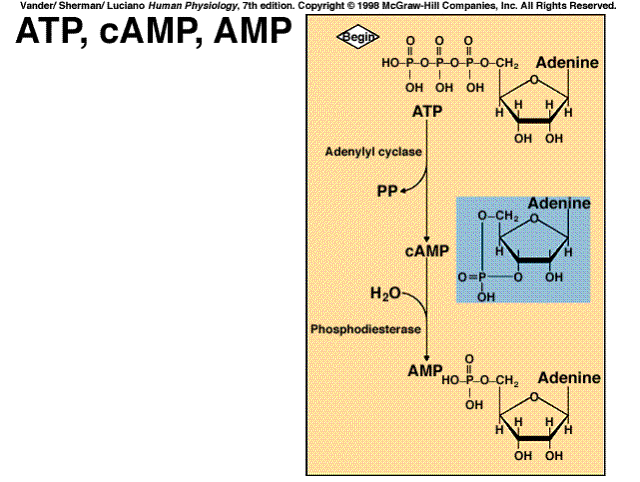 |
+effects: Activates cAMP-dependent protein kinase (protein kinase A)
|
|
 |
+source: Generated from guanosine triphosphate in a reaction catalyzed by a plasma-membrane receptor with guanylyl cyclase activity
+effects: Activates cGMP-dependent protein kinase (protein kinase G)
+source: A G protein activates plasma-membrane
phospholipase C, which catalyzes generation of DAG and IP3 from
plasma membrane phosphadidylinositol bisphosphate (PIP2)
+effects: Activates protein kinase C
+source: See DAG above
+effects: Releases calcium from endoplasmic reticulum
===================================================
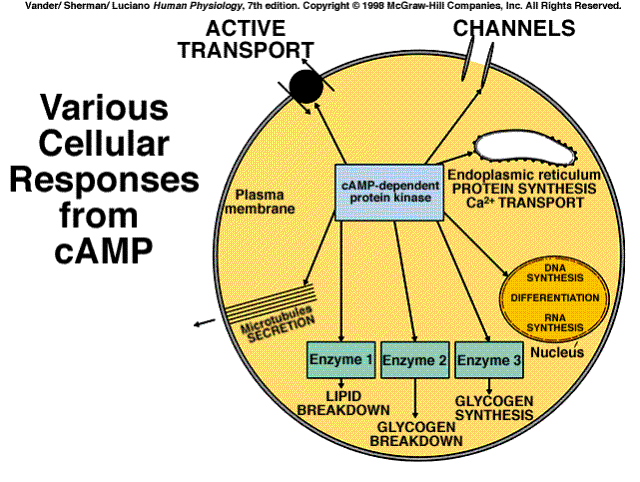
The cyclic AMP system is a classic example of a cascade amplification system. Each stage of the system can amplify the effects hundreds of times. The net result is that one messenger molecule can cause the production of a million products.
|
|
 |
===================================================
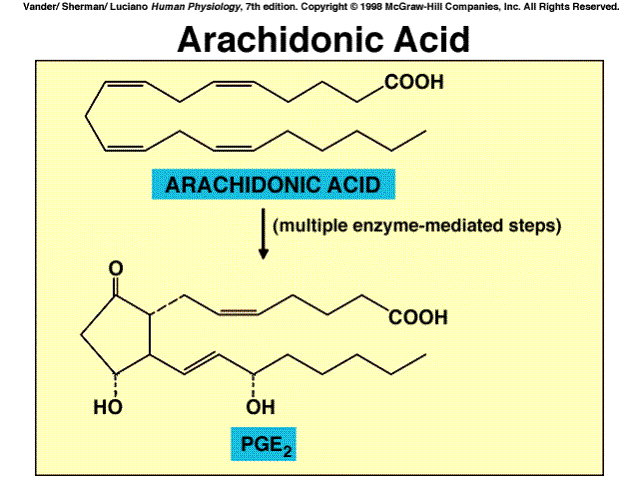
One set of paracrine/autocrine agents which exerts a wide variety of effects
in virtually every tissue and organ system are the eicosanoids. These are
a family of substances produced from the polyunsaturated fatty acid arachidonic
acid.
|
|
 |
They include the cyclic endoperoxides, the prostaglandins, the thromboxanes, and the leukotrienes.
|
|
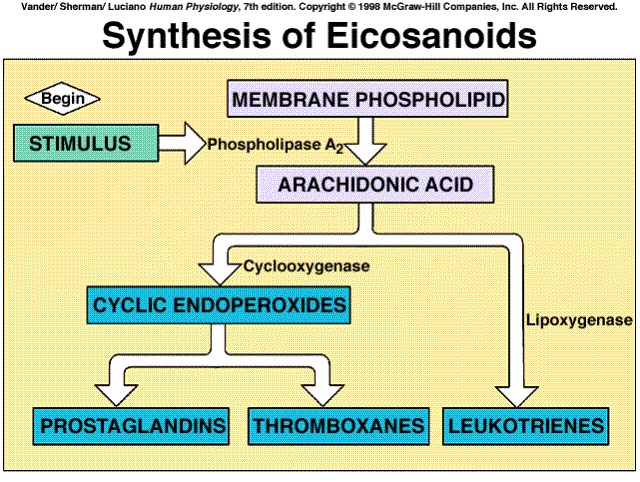 |
Drugs that influence the eicosanoid pathway include Aspirin, which inhibits cyclooxygenase and, therefore, blocks the synthesis of the endoperoxides, prostaglandins, and thromboxanes. It, and a variety of new drugs that also block cyclooxygenase, are collectively termed nonsteroidal anti-inflammatory drugs (NSAIDs). Their major uses are to reduce pain, fever, and inflammation. The term "nonsteroidal" distinguishes them from the adrenal steroids that are used in large doses as intiinflammatory drugs; these steroids inhibit phospholipise A2 and thus block the production of all eicosanoids.
Hormones are the technical name for chemical messangers utilized by the endocrine system. While these are frequently the same chemical messengers utilized by other systems, when they are used by endocrine glands they are distinguished by being called hormones.
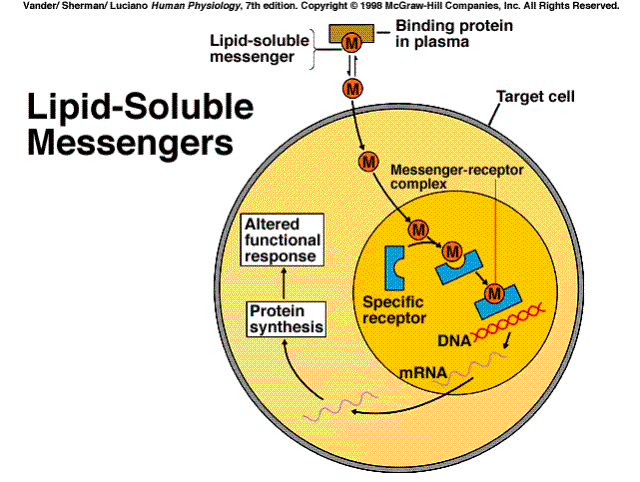
In Part III, Characteristics of Messangers, we talked about membrane soluble and insoluble messengers. Hormones are also either lipid soluble or insoluble. The lipid insoluble receptors use the mechanisms previously delineated. The lipid soluble messengers enter the cell and initiate responses. The messenger may complex either with receptors located in the cytoplasm or the nucleus. When the messenger complexes with a receptor in the cytoplasm, it subsequently passes into the nucleus. The messengers which pass directly into the nucleus, complexes with receptors in the nucleus, and initiates an action, e.g., protein synthesis by the transcription of mRNA.
|
|
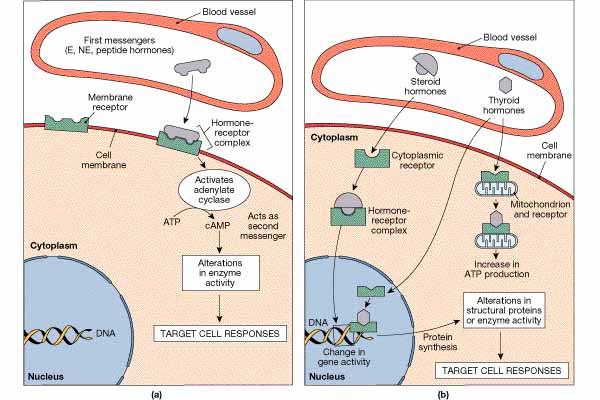 |
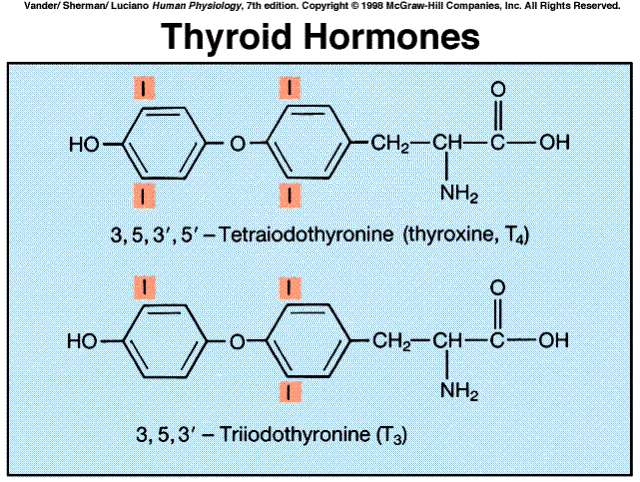
The amine hormones are all deriveatives of the amino acid tyrosine. They include the thyroid hormones (produced by the thyroid), epinephrine and norepinephrine (produced by the adrenal medulla), and dopamine (produced by the hypothalamus).
|
|
 |
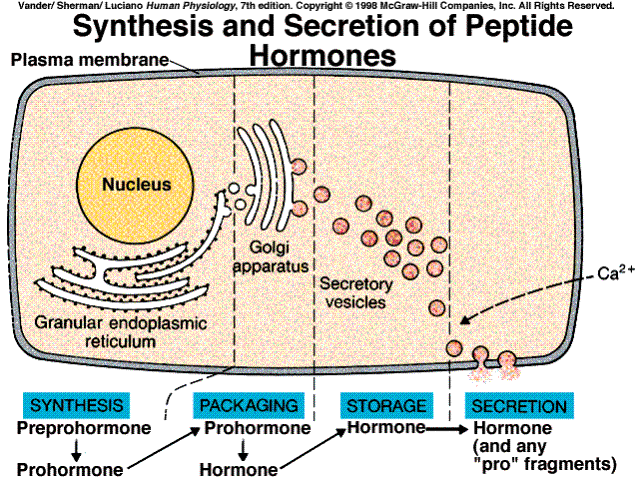
The great majority of hormones are either peptides or proteins. they range in size from small peptides having only three amino acids to small proteins (some of which are glycoproteins). In many cases, they are initially synthesized on the ribosomes of the endocrine cells as larger proteins known as preprohormones, which are then cleaved to prohormones by proteolytic enzymes in the granular endoplasmic reticulum. The prohormone is hen packaged into secretory vesicles by the Golgi apparatus. In this process, the prohormone is cleaved to y6eald the active hormone and other peptide chains found in the prohormone.
When the cell is stimulated to release the contents of the secretory vesicles by exocytosis, the other peptides are cosecreted with the hormone. Instead of just one peptide hormone, the cell may be secretihg multiple peptide hormones that differ in their effects on target cells.
|
|
 |
Steroid hormones are produced by the adrenal cortex and the gonads (testes and ovaries), in the placenta during pregnacy, and in the skin, Vitamin D.
|
|
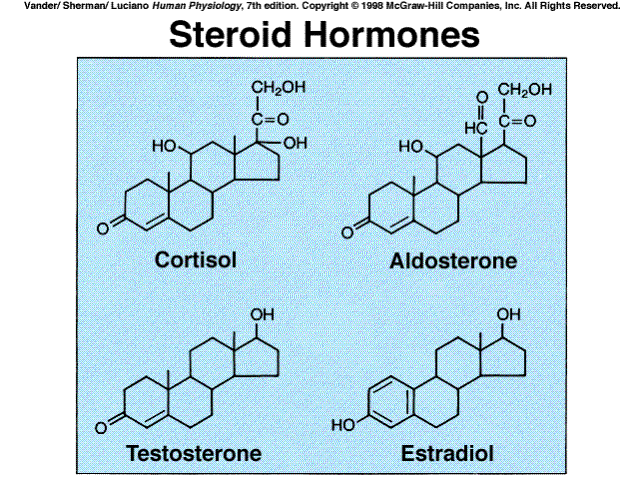 |
Cholesterol is the precursor of all steroid hormones. The cells producing these hormones synthesize some of their own cholesterol, but most is provided to them from the plasma. The many biochemical steps in steroid synthesis beyond cholesterol involve small changes in the molecules and are mediated by specific enzymes. Because steroids are highly lipid-soluble, once they are synthesized, byey simply diffuse across the plasma membrane of the steroid-producing cell and enter the interstitial fluid and then the blood where they mainly bind to certain plasma proteins.
|
|
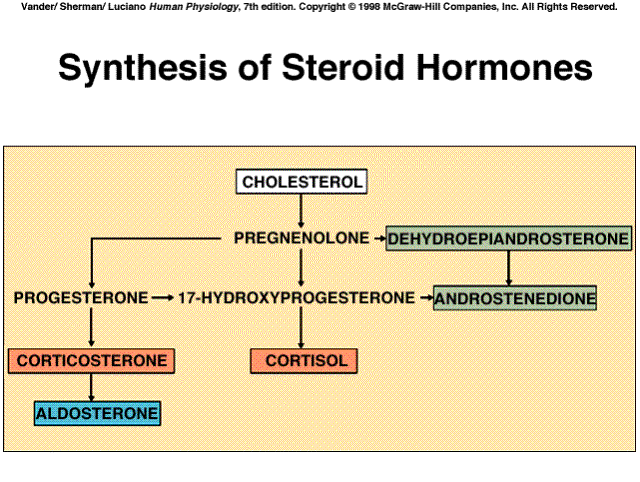 |
Several intermediate steps have been left out of the diagram. The five
hormones shown in boxes are the major hormones secreted.
|
|
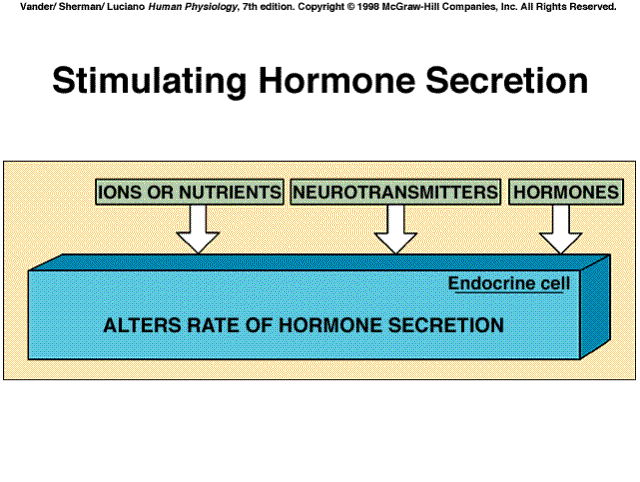 |
Most hormones are released in short bursts, with little or no release occurring between bursts. Therefore, the plasma concentrations of hormones may fluctuate rapidly over brief time periods. Hormones also manifest circadian, 24-hour, cyclical rythms in their secretory rates. the circadian patterns are different for different hormones. some are clearly linked to sleep, e.g., growth hormone. The mechanisms underlying these cycles are ultimately traceable to cyclical variations in the activity of neural pathways involved in the hormone's release.
Hormone secretion is controlled by three principal types of inputs to the endocrine cells. These are changes in the plasma concentrations of mineral ions or organic nutrients. Neurotransmitters released from neurons impinging on the endocrine cell. Other hormones, or paracrine/autocrine agents acting on the endocrine cell. In many cases, hormone secretion is influenced by more than one input. Thus, endocrine cells, like neurons, may be subject to multiple, simultaneours, often opposing inputs, and the resulting output reflects the integration of all these inputs.
Hormones can influence the ability of target cells to respond by regulating hormone receptors. In the context of hormones, up-regulation is an increase in the number of a hormone's receptors, often resulting from a prolonged exposure to a low concentration of the hormone. Down-regulation is a decrease in receptor number, often from exposure to high concentrations of the hormone.
Hormones can down-regulate or up-regulate not only their own receptors but the receptors for other hormones as well. If one hormone induces a loss of a second hormone's receptors, the result will be a reduction of the second hormone's effectiveness. In such cases the one hormone is said to antagonize the action of the other. Alternatively, a hormone may induce an increase in the number of receptors for a second hormone. In this case the effectiveness of the second hormone is increased.
This latter phenomenon underlies the important hormone-hormone interaction known as permissiveness. In general terms, permissiveness means that hormone A must be present for the full strength of hormone B's effect. A low concentration of hormone A is usually all that is needed for this permissive effect, which is due to A's positive effect on B's receptors. For example, epinephrine causes a large release of fatty acids from adipose tissue, but only in the presence of permissive amounts of thyroid hormone. The major reason is that thyroid hormone facilitates the synthesis of receptors for epinephrine in adipose tissue and so the tissue becomes much more sensitive to epinephrine.
The events initiated by the binding of a hormone to its receptor are one or more of the signal transduction pathways that apply to all chemical messengers. That is, the mechanisms initiated by hormones are the same as those utilized by neurotransmitters, paracrine, and autocrine agents.
|
|
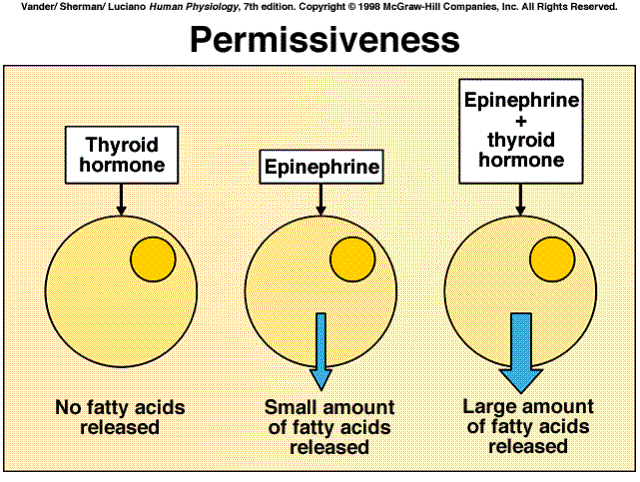 |
A hormone's concentration in the plasma depends both on its rate of secretion and upon its rate of removal from the blood. Removal may be by excretion or by metabolic transformation. The liver and the kidneys are the major organs that excrete or metabolize hormones.
Hormones are also metabolized by the cells upon which it acts. Endocytosis of hormone-receptor complexes on plasma membranes enables cells to remove the hormones rapidly from their surfaces and catabolize them intracellularly. the receptors are then often recycled to the plasma membrane.
Catecholamine and peptide hormones are excreted rapidly and are attacked by enzymes in the blood and tissue and remain in the bloodstream for only brief periods, minutes. Protein-bound hormones are less vulnerable and circulating steroid and thyroid hormones takes much longer, hours to days.
Metabolism of the hormone after secretion may activete the hormone, rather than inactivate it. Thyroid T4 is converted to the far more active T3. Testosterone is converted to estradiol or dihydrotestosterone in certain of its target cells. It is these molecules, rather than testosterone, which bind to the receptors inside the target cell and elicit the cell's response.
Sometimes, instead of the hormone itself being activated after secretion, it actually acts enzymatically on a completely different plasma protein to split off a peptide that functions as the active hormone. In the renin-angiotensis system, renin acts as an enzyme that participates in the generation of the hormone angiotensin.
|
|
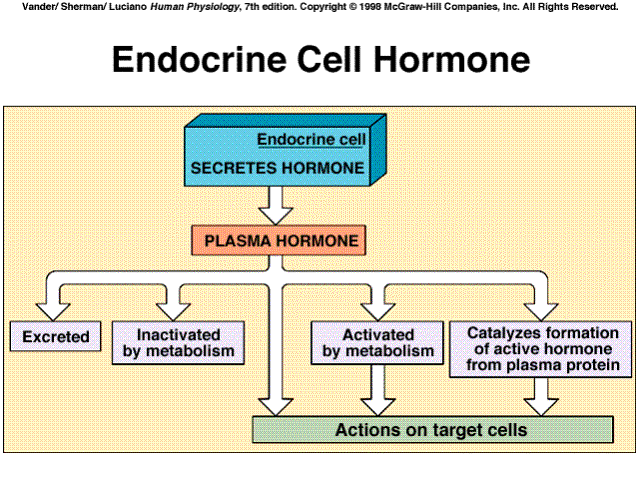 |
